

Por coevolución negativa entendemos todas los procesos de coevolución que conllevan a una situación negativa para al menos una de las especies involucradas. Se incluyen la depredación, el parasitismo y la competencia. Veamos cada uno de ellos con un poco más de detalle.
La depredación y el parasitismo se pueden considerar como dos caras de la misma moneda, en la que una especie se alimenta a costa de otra. Esto puede llevar al consumo total del individuo como ocurre en la depredación, o parcial como en el parasitismo. La distinción no es clara desde una perspectiva práctica porque muchos casos de depredación no acaban con la vida del organismo. Por ejemplo, la mayoría de los herbívoros no consumen toda la presa. Por otra parte, algunos casos de parasitación sí que lo hacen, como la enfermedad de la malaria (paludismo), causada por un parásito (protista), que puede acabar matando a muchas personas. Tanto los depredadores/parásitos como las presas/huéspedes han desarrollado extraordinarias modificaciones evolutivas (anatómicas, fisiológicas, de comportamiento, etc) para tratar de mejorar sus interacciones con la otra parte. Así, por ejemplo, las presas/huéspedes han mejorado su sistema inmunitario o desarrollado diferentes estrategias de escape, y los parásitos/depredadores han evolucionado para poder eludir esas estrategias de defensa. Nos vamos a centrar en la relación depredador/presa, pero desde una perspectiva teórica es idéntica a la relación parásito/presa o herbívoro/planta. En teoría la coevolución entre un depredador y su presa puede evolucionar siguiendo alguno de los siguientes escenarios alternativos:
1. Una de las especies se puede extinguir. Esta situación, aunque fácil de entender y modelar es, por lo general, poco frecuente. Se puede observar, por ejemplo, con la introducción de nuevas especies en un determinado hábitat (especies invasoras) o en situaciones de estrés ambiental (contaminación, degradación de hábitat, etc). En la mayoría de los casos es más frecuente observar las dos alternativas siguientes.
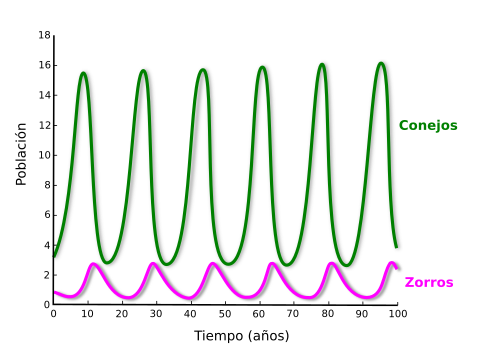
2. Pueden producirse ciclos continuos o discontinuos favoreciendo al depredador o la presa. Depredador y presa suelen estar demográficamente relacionados, pues una población sana de presas redunda en beneficio del depredador, aunque un exceso en el número de depredadores acaba ocasionando una reducción en la densidad de la presa, lo que a la larga de nuevo acaba afectando a la población de depredadores. Esta relación se puede modelar matemáticamente por una serie de ecuaciones diferenciales características de la ecología (Figura 1).
3. Se alcanza una situación de equilibrio evolutivo dinámico (hipótesis de reina roja; Van Valen 1973). El equilibrio anterior se refiere a la relación demográfica en el tiempo actual, pero esta explicación de equilibrio evolutivo hace referencia a un equilibrio a muy largo plazo. La idea es que la especie de presa evoluciona en el sentido de mejorar sus características de escape respecto al depredador, con lo que con el tiempo el depredador tendrá que evolucionar de forma concomitante a la evolución de su presa si quiere mantener su eficacia depredadora. Con el tiempo se produce una carrera evolutiva en la que las diferencias entre depredador y presa se mantienen aproximadamente constantes. Esta hipótesis representa una poderosa metáfora de cómo pueden interaccionar depredador y presa evolutivamente, aunque es difícil de probar experimentalmente o incluso modelar teóricamente.
Un ejemplo de coevolución depredador/presa lo tenemos en la relación que hay con un linaje de peces depredadores y sus presas de moluscos gasterópodos. Los peces de la familia Tetraodontidae, vulgarmente conocidos como peces globos, representan un linaje en el que como respuesta a las conchas que protegen a muchas de sus presas han desarrollado un aparato bucal especializado en el aplastamiento de caracoles. Como respuesta evolutiva a este y otros tipos similares de depredadores un grupo de gasterópodos marinos han evolucionado presentando una serie de espículas finas y largas que alejan la parte sensible de la concha de las peligrosas mandíbulas de estos depredadores (Palmer 1979; Figura 2).

{kind=link}
{kind=link}
Evolución competitiva
Otro tipo de interacción evolutiva negativa entre especies afectadas por la selección natural es la evolución competitiva. La evolución competitiva se da cuando dos o más especies coinciden total o parcialmente en cierto hábitat y comparten ciertos recursos finitos (espacio, agua, alimentos, etc). De esa forma la habilidad de cada especie para aprovechar dicho recurso va a estar condicionada por la presencia de la otra especie, y de alguna forma se puede desatar una carrera evolutiva entre ambos linajes por aprovechar mejor (o diferentemente) los recursos en disputa. Algunos autores han descrito, desde una perspectiva ecológica, que las especies que compiten para un determinado recurso importante, con el tiempo, o bien una se extingue o bien una o ambas evolucionan hacia un uso diferente del recurso en disputa. Es lo que se denomina el principio de exclusión competitiva (Hardin 1960). Este principio está en discusión hoy en día, pues no está del todo claro si siempre se tiene que dar o no. Lo que sí parece que ocurre “habitualmente” es la tendencia evolutiva de dos especies que compiten por un recurso a evolucionar de forma diferente, minimizando de esa forma su competencia, lo que suele denominarse como desplazamiento de carácter (Brown y Wilson 1956). El desplazamiento de carácter, se observa frecuentemente en especies que mantienen poblaciones alopátridas (viven en territorios diferentes) y simpátridas (comparten el mismo territorio), de forma que en las que conviven en simpatría, se pueden observar cambios en el comportamiento o la morfología que van encaminados a minimizar la competición entre las especies. Un buen ejemplo de esto lo tenemos en las especies de pinzón de las islas galápagos Geospiza fortis y G. fuliginosa (explicado en Futuyma 2009, páginas 516-517). En promedio, G. fortis presenta anchuras de pico ligeramente mayores que G. fuliginosa, como consecuencia de que se alimentan de semillas más grandes, aunque muchas de las semillas de las que se alimentan caen en un rango de tamaño que ambas especies pueden utilizar, y para las que compiten. Así los Grant (un matrimonio que trabajaban juntos como científicos) demostraron que los tamaños de pico de las especies anteriores divergían más entre sí cuando ambas especies conviven en simpatría (isla de Santa Cruz) que cuando viven en islas separadas (Figura 3; Futuyma 2009).

Para saber más:
Brown, W.L. y Wilson, E.O. 1956. Character displacement. Systematic Zoology 5: 49–64. Futuyma, D. 2009. Evolution, second edition. Sinauer Associates, Inc., Massachusetts.
Hardin, H. 1960. The competitive exclusion principle. Science 131: 1292–1297.
Palmer, A.R. 1979. Fish predation and the evolution of gastropod shell sculpture: experimental and geographic evidence. Evolution 33: 697-713.
Van Valen, L. 1973. A new evolutionary law. Evolutionary Theory 1: 1-30.