


Existen diversos métodos de estimación de los efectos de selección natural para un carácter cualitativo (ver ejemplos en Cook et al. 1999). Como ya hemos remarcado con anterioridad nos centraremos en los diseños transversales (ver apartado anterior), es decir, vamos a tratar de estimar la eficacia biológica de caracteres cuantitativos comparando el cambio en frecuencia entre dos momentos de una población, y antes y después de que haya actuado la selección. Estos momentos pueden ser generaciones distintas (sería una estimación de la eficacia total) o diferentes momentos del ciclo de vida de los organismos (componentes de viabilidad, selección sexual, fecundidad, etc).
De entre los diferentes métodos que se han propuesto para medir la eficacia en caracteres cualitativos vamos a explicar el método de la fracción de morfos (variantes de un fenotipo; Cook et al. 1999), también denominado el estimador de productos cruzados (descrito habitualmente con la letra “W”). Para explicar el método lo más sencillo es partir de un hipotético ejemplo de carácter cualitativo estudiado en una especie en dos momentos del ciclo de vida (cuando los ejemplares son juveniles y cuando son adultos; ver Tabla 1).
| Littorina saxatilis | Clara | Oscura |
| Juveniles | 123 | 133 |
| Adultos | 34 | 67 |
El método de la fracción se inicia con el cálculo de la fracción fD/fA (frecuencia del carácter DESPUES / frecuencia del carácter ANTES, de que actúe la selección). Para Claro sería 34/123=0.28 y para Oscuro sería 67/133=0,50. Se ha comprobado que estadísticamente tiene ventajas presentar este valor relativo de un morfo respecto a otro (simplemente se dividen todas las fracciones por el valor de una de ellas). Por ejemplo si queremos utilizar el color claro como referencia, dividimos las fracciones anteriores por el valor de claro, esto sería para claro 0,28/0,28=1 y para oscuro 0,50/0,28=1.8. La interpretación de estos valores es directa, los ejemplares oscuros en esta población parecen tener una ventaja en su eficacia biológica de 1,8 respecto a 1, o lo que es lo mismo su eficacia es un 56% mayor (1/1,8 × 100). Una propiedad de este estimador es que se puede relativizar a diferentes morfos, por ejemplo podríamos haber usado el color oscuro como referencia en cuyo caso las eficacias habrían sido W=0,28/0,50=0,56 para el claro y W=0,50/0,50=1 para el oscuro. Nótese que las diferencias relativas de eficacia son las mismas por ambos métodos si la expresamos en porcentaje, utilizando el claro como referencia la eficacia del oscuro es un 56% mayor, mientras que si usamos el oscuro como referencia la eficacia del claro será un 56% menor. La interpretación en ambos casos es la misma: los ejemplares oscuros en esta población y en este periodo estudiado fueron mayores en los oscuros que en los amarillos.
Cómo estamos midiendo eficacias biológicas, y como asumimos que los caracteres estudiados (salvo que se indique lo contrario) tienen una base genética, la consecuencia esperada de esta situación sería (previsiblemente) un aumento de la frecuencia del color oscuro en la generación siguiente, aumento que en ciertas condiciones se puede comprobar. La realidad es un poco más compleja. Para empezar, para poder hacer predicciones precisamos conocer en detalle la herencia genética del color de la concha, si está causada por uno o varios genes, o si presenta otras complicaciones (epistasias -cuando los alelos de dos genes interaccionan de forma no aditiva para producir el fenotipo-, influencia del ambiente, etc). Además, para cualquier intento predictivo hay que asumir otra serie de simplificaciones, si es que no se pueden comprobar directamente en el campo, y estas serían:
1. Que la eficacia estimada es estable en el tiempo. Es decir, que no cambia de un mes a otro, de un año a otro, etc.
2. Que el cambio de frecuencia causado por la eficacia va a ser más importante que el causado por cualquier otra fuerza evolutiva (migración, mutación o deriva genética).
3. Que el cambio de frecuencia observado no pueda ser explicado por procesos fortuitos durante el muestreo (al igual que tirando repetidamente una moneda podemos observar desviaciones de las proporciones esperadas: 50% cara y 50% cruz). Este aspecto es importante, aunque se sale del nivel del curso diseñado en estos capítulos, pero cualquier estudio empírico sobre la estimación de la eficacia requiere de algún test estadístico que rechace la posibilidad de que las observaciones puedan explicarse por azar. En este caso se podría aplicar un test chi-cuadrado sobre los datos de la tabla 1. Cualquier lector interesado en los test estadísticos que se podrían aplicar a datos de este tipo puede consultar el Sokal y Rohlf (1995).
Si se dan estas circunstancias, podríamos inferir que en la población estudiada se producirá con el tiempo un aumento gradual pero continuado del carácter oscuro, pudiendo llegar la población a convertirse en monomórfica (con un solo color).
Vamos a poner un ejemplo reciente a continuación del uso del estimador W (fracción de morfos; ver Estévez et al. 2021). Estos autores estudiaron ejemplares de la especie L. fabalis en la población de Abelleira (Ría de Muros-Noya, Galicia, España) durante 7 años, aunque se conocen estimas de la misma población desde hace más de 20 años. Estudiaron el carácter patrón de bandeado en la concha, pues en dicha población aproximadamente un 10% de los caracoles presentan un patrón visible de bandeado (Figura 1), que aparentemente ha permanecido constante durante más de 20 años, sugiriendo algún mecanismo selectivo (como selección dependiente de las frecuencias; SDF) manteniendo el polimorfismo. Para verificar la hipótesis de SDF se estimaron las eficacias para el carácter bandeado utilizando las frecuencias de años consecutivos (ver tabla 2). Como se observa las estimas alternaron entre valores a favor y en contra del carácter de forma correlacionada con su frecuencia como se puede observar en la Figura 2.
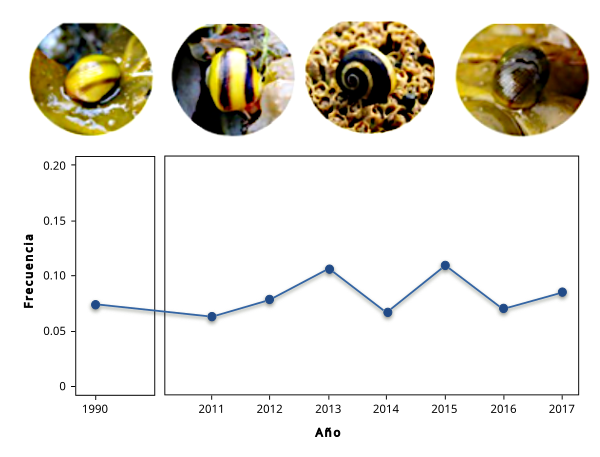
| Intervalo de años | N | % Bandeados | W |
| 2011-2012 | 521 | 6.3 | 1.27 |
| 2012-2013 | 708 | 7.9 | 1.38* |
| 2013-2014 | 574 | 10.6 | 0,61* |
| 2014-2015 | 564 | 6.7 | 1.70* |
| 2015-2016 | 593 | 11.0 | 0.61* |
| 2016-2017 | 841 | 7.0 | 1.23 |
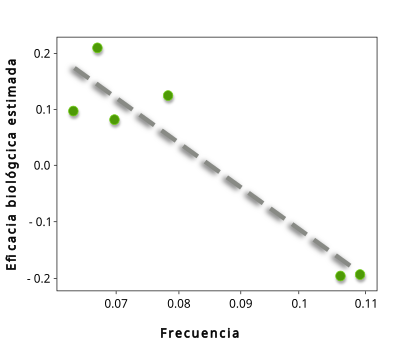
En este estudio no se identificó el mecanismo biológico causante de esta selección dependiente de las frecuencias, pero en éste y otros estudios similares en gasterópodos se ha sugerido que podría ser causado por selección causada por alguno de sus depredadores más frecuentes (como peces de la familia Blenniidae). A este tipo de selección se le denomina selección apostática. La selección apostática ocurre porque el depredador tiende a fijarse en el morfo más frecuente, de forma que cuando la frecuencia se hace muy baja para cierto morfo, el depredador cambia la percepción de cuál es el morfo más interesante empezando a fijarse en el más frecuente, y por lo tanto la eficacia del morfo a baja frecuencia aumenta.
Para saber más:
Cook, L.M., Cowie, R.H. and J.S. Jones. 1999. Change in morph frequency in the snail Cepaea nemoralis on the Marlborough Downs. Heredity, 82: 336-342.
Estévez, D., Galindo, J. y E. Rolán-Alvarez. 2021. Negative frequency dependent selection maintains shell banding polymorphisms in two marine snails (Littorina fabalis and L. saxatilis). Ecology and Evolution 11: 6381-6390.
Sokal, R.R. y F.J. Rohlf. 1995. Biometry. Freeman and Co., New York.