

Darwin fue el primero en proponer un mecanismo de selección que desde su punto de vista era diferente al de la selección natural. Lo propuso para explicar la existencia de ciertos caracteres extravagantes que se observan en uno de los sexos en diversas especies (Futuyma 2009). Por ejemplo, los machos de muchas aves presentan unos caracteres de forma, color y tamaño de sus colas tan extravagantes que difícilmente podían haber evolucionado por selección natural exclusivamente debido a que al portador de dicho carácter le acarrea un problema a la hora de escapar de posibles depredadores (Figura 1). Según Darwin estos caracteres se explican por la existencia de dos nuevos mecanismos que, más que explicar la lucha por la existencia, explican la lucha por conseguir aparearse (una parte relevante del éxito reproductivo total): la competencia dentro de sexos y la elección de uno de los sexos por el otro. Es decir, según Darwin, estos caracteres extravagantes se desarrollaban en ciertos machos porque, a pesar de que pudieran ser contraproducentes para la supervivencia del macho, le conferían a estos grandes ventajas a la hora de alcanzar la fecundación de las hembras de su propia especie, y por tanto aumentar la probabilidad de dejar más descendencia. Estas ventajas podrían ser ganar en las competiciones con otros machos, o bien ser elegidos preferentemente por las hembras. Estos mecanismos de competencia y elección definirían lo que se conoce como la selección sexual, la cual representa la contribución de dichos mecanismos a la eficacia biológica.

{kind=link}
La importancia relativa de los mecanismos que determinan la selección sexual, y considerarlos o no como un proceso realmente diferente de la selección natural, han sido fuente de múltiples discusiones. Actualmente ambas estrategias (considerar o no a la selección sexual como diferente de la selección natural) tienen partidarios: por un lado, hay autores que sostienen que la selección sexual es diferente a la selección natural y otros que postulan que la selección sexual es un componente más de la selección natural (ver defensas de ambos puntos de vista en Henshaw et al. 2018; Hosken y House 2011). En este curso nosotros usaremos la interpretación más sencilla, que es que la selección sexual es sólo un componente más en los que se puede dividir la selección natural, y que no hay conceptualmente nada en ella que la distinga y la separe de la selección natural (ver capítulo 1 de este tema). No obstante la selección sexual, aún siendo un componente de la selección natural, tiene algunas peculiaridades que conviene tener presentes cuando se trata de medirla o interpretarla. La primera peculiaridad es que conviene estimar el componente de la selección sexual por separado para machos y hembras, pues es relativamente frecuente que la fuerza de la selección sexual difiera entre sexos de la misma especie.
Más peculiaridades de la selección sexual las podemos ver mediante la estima de caracteres cualitativos y cuantitativos en un ejemplo experimental real. Por ejemplo, unos investigadores realizaron una serie de experimentos para tratar de entender las fuerzas que modelan la evolución de un carácter: el tamaño de la concha en la especie Echinolittorina malaccana en Hong Kong (Lau et al. 2021). Para ello, subdividieron el carácter tamaño de la concha en 4 clases cualitativas T1, T2, T3 y T4, siendo T1 la clase más pequeña y T4 la más grande (de igual forma en machos y hembras). Cuatro machos y cuatro hembras, un representante de cada clase de tamaño, se dispusieron juntos en unas cámaras de apareamiento (unas bolas plásticas ver Figura 2), de forma que se pudo anotar cuales de los ejemplares finalmente se apareaban y cuales no, en función de su tamaño y sexo. Este experimento se repitió 23 veces. Un resumen simplificado de sus resultados aparece en la Tabla 1.
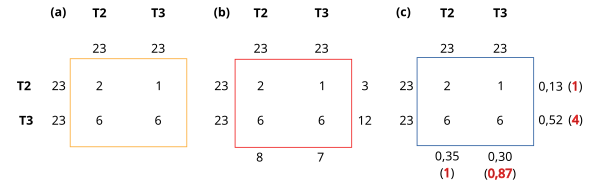
Como se explica en la leyenda de la propia tabla, cada una de las versiones de la tabla 1 (a-c) son los diferentes pasos que se dan para realizar los cálculos a partir de los datos acumulados en la tabla de apareamientos (Tabla 1a). A diferencia de lo que se hace en estudios con eficacias no sexuales, en este caso tenemos los datos de fuera de la tabla (izquierda y arriba) que representan los ejemplares utilizados en el experimento para los diferentes tamaños, y tenemos los datos de dentro del recuadro que representan un tipo de pareja cada uno. Por lo tanto habrá que contabilizar de forma independiente para machos y para hembras (Tabla 1b). Finalmente en la Tabla 1c se presentan los cálculos para obtener el estimador W, que es la razón entre la frecuencia del carácter después de que ha actuado la selección (sexual en este caso) y antes de la selección, relativizada a uno de los caracteres (ver tema estimación de las eficacias en caracteres cualitativos).
La interpretación de los resultados es bastante intuitiva: en machos prácticamente no hay diferencias en las eficacias sexuales para las clases T2 y T3 (1 vs 0.87), mientras que las hembras más grandes (T3) presentan una eficacia sexual 4 veces más grande que las más pequeñas (T2). Este efecto de selección sexual se produce porque, como demuestran estos autores, los machos de esta especie presentan una marcada preferencia durante los apareamientos por hembras que son un 10% mayores que ellos mismos, y por eso la clase mayor en hembras acaba apareciendo beneficiada. Si no hubiese otros factores evolutivos actuando en esta población, esta fuerza favorecería un incremento del tamaño en la especie o cuanto menos en las hembras (esto último, por otra parte, que sí es observado en la naturaleza).
El mismo ejemplo puede usarse para estudiar un estimador de selección sexual en un carácter cuantitativo. En un trabajo anterior, Ng y coautores (2019), se midieron los tamaños medios de machos y hembras de E. malaccana que estaban apareándose (en cópula) y de aquellos que estaban cerca de tales apareamientos. Como puede verse en la tabla 2, el estimador diferencial de selección (S) puede fácilmente obtenerse al sustraer el valor medio del carácter (tamaño) en los ejemplares no apareados del valor de los ejemplares apareados, siendo claramente superior en hembras. En la tabla 2 se muestran los valores de S en bruto, pero normalmente se presenta dichos valores estadarizados, es decir, se dividen dichos valores por la desviación típica del carácter en cada sexo.
| Sexo | Apareados | No apareados | S |
| Machos | 8.44±1.38 | 8.11±1.44 | 0.33 |
| Hembras | 9.23±1.45 | 8.53±1.58 | 0.7 |
Para saber más:
Futuyma, D. 2009. Evolution. Second Edition., Sinauer associates, inc., Massachusetts.
Henshaw, J., Jennions, M. y L. Kruuk. 2018. How to quantify (the response to) sexual selection traits. Evolution, 72: 1904-1917.
Hosken, D.J., y C.M. House. 2011. Sexual selection. Curr Biol., 21(2): R62-5.
Lau, S.L.Y., Williams, G.A., Carvajal-Rodríguez, A., y E. Rolán-Alvarez. 2021. An integrated approach to infer the mechanisms of mate choice for size. Animal Behaviour, 175: 33-43.
Henshaw, J., Jennions, M. y L. Kruuk. 2018. How to quantify (the response to) sexual selection traits. Evolution, 72: 1904-1917.
Ng, T.P.T., Rolán-Alvarez, E., Dahlén, S.S., Davies, M.S., Estévez, D., Stafford, R., y G.A. Williams. 2019. The causal relationship between sexual selection and sexual size dimorphism in marine gastropods. Animal Behaviour, 148: 53-62.