

La selección natural (de aquí en adelante usaremos simplemente selección para abreviar) consiste en la reproducción diferencial no aleatoria de los organismos de una población en un determinado ambiente, que beneficia sistemáticamente a aquellos organismos que poseen genes que les proporcionan una mayor supervivencia y reproducción. La selección natural es el principal mecanismo evolutivo postulado por la teoría de la evolución, pues solo ella permite explicar de forma satisfactoria la aparición de las adaptaciones.
De acuerdo con la formulación clásica del biólogo evolutivo Richard Lewontin (1970), para que actúe la selección en una población es necesario que se cumplan tres condiciones:
1) Que exista variación fenotípica en una población, es decir, que la anatomía, fisiología y conducta de los distintos miembros de la población no sea idéntica.
2) Que la variación fenotípica que existe en la población sea heredable entre padres e hijos, de forma que estos últimos se parezcan a los primeros en mayor medida que a cualquier otro miembro aleatorio de la población.
3) Que los distintos fenotipos de dicha población tengan diferente habilidad para sobrevivir y reproducirse en un ambiente determinado. En otras palabras, que la variación fenotípica presente en la población conlleve variación en eficacia reproductiva (fitness ).
En la medida en que se den estas tres condiciones existirá selección en una población.
Habitualmente se admite que la selección puede actuar sobre los distintos niveles de organización biológica (los genes, los individuos, los grupos, etc.); sin embargo, dado que para la teoría evolutiva estándar (la Síntesis Evolutiva Moderna; SEM) la herencia fenotípica depende en grado sumo de los genes que porta un individuo, se entiende que el proceso selectivo más importante es aquel que tiene lugar sobre ellos. Según el modelo estándar, los genes permiten que exista variación fenotípica en las poblaciones (a través de las mutaciones o la recombinación genética), que existan diferencias en la habilidad para sobrevivir y reproducirse de cada organismo (debido a la diferente fitness que lleva asociada cada alelo) y que la variación fenotípica se herede a la descendencia (a través de la transmisión de los genes entre generaciones). Así pues, para la teoría evolutiva moderna los genes y su estudio adquieren una importancia capital para entender y comprender el proceso evolutivo.
Tipos de selección natural
Habitualmente se admite que la selección puede actuar sobre los distintos niveles de organización biológica (los En función del tipo de cambio evolutivo que produce sobre una población, es posible distinguir cuatro tipos de selección: direccional, estabilizadora, disruptiva y equilibradoras. Para entender cómo funciona cada uno de estos cuatro tipos, imaginemos un rasgo fenotípico determinado que se distribuye en una población siguiendo una distribución normal o campana de Gauss (Figura 1). Así, en el eje X del gráfico se representan los valores que puede adoptar dicho rasgo, mientras que en el eje Y se representa el número de individuos de la población que presentan cada valor determinado del rasgo.

Pues bien, lo que sucede en la selección direccional es que la media de los valores del rasgo analizado en la población se desplaza en una dirección determinada, pasando a ser mayor o menor que la que existía antes del proceso de selección (Figura 2). El motivo de este desplazamiento es que la selección natural actúa en contra de aquellos individuos que presentan valores extremos en un sentido y a favor de aquellos individuos que presentan valores extremos en el sentido opuesto.

Un ejemplo de selección direccional lo constituye el caso de los picos de los pinzones de las islas Galápagos estudiados por Peter y Rosemary Grant, que comentamos en el apartado sobre “Evolución en el laboratorio y en la naturaleza”. En él, tras la gran sequía que se produjo en el año 1977, que eliminó del ecosistema las semillas de tamaño medio, se produjo un proceso de selección direccional que favoreció a aquellos individuos de la población con una altura de pico más grande, dado que solo estos eran capaces de aprovechar las semillas de gran tamaño que sobrevivieron a la sequía. Como resultado, la siguiente generación de pinzones mostró un promedio de altura de pico mayor que la de generaciones anteriores.
Por otro lado, en la selección estabilizadora el proceso de selección perjudica a los extremos de la distribución, aumentando la frecuencia de casos con valores centrales que estén cercanos al promedio (Figura 3). A diferencia de la selección direccional, la selección estabilizadora provoca la estasis de la población, es decir, que con el tiempo la población se mantiene con la misma media. De ese modo, la selección estabilizadora contribuye a reducir o mantener a raya la cantidad de variación genética y fenotípica de la población.

En tercer lugar, la selección disruptiva es aquella en la que los extremos obtienen una ventaja, mientras que los valores centrales salen perjudicados. Aquí ocurre exactamente el proceso contrario al que tiene lugar en la selección estabilizadora que vimos anteriormente (Figura 4).
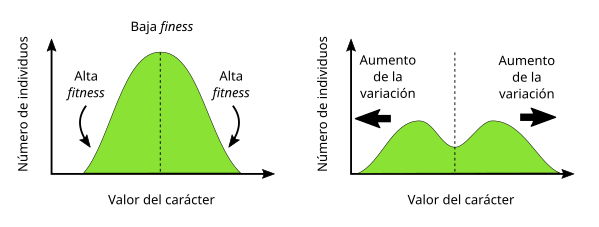
La selección disruptiva contribuye a mantener la diversidad genética de la población al favorecer los alelos responsables de los valores extremos de la distribución del rasgo. Además, este tipo de selección es importante en la medida en que puede ser el inicio de un proceso de especiación, al contribuir a la diferenciación de la población en dos subpoblaciones con valores muy distintos para un rasgo en concreto, algo que más adelante veremos cómo puede contribuir a la emergencia de un aislamiento reproductivo asociado a dicha diferenciación.
Un ejemplo particularmente ilustrativo de selección disruptiva se ha visto en el pinzón cascanueces de vientre negro (Pyrenestes ostrinus; Figura 5), cuyas poblaciones presentan individuos con longitudes de pico muy distintas (o bien muy cortos, o bien muy largos) (Smith, 1987). La investigación ha mostrado que los individuos con un tamaño de pico intermedio se ven penalizados por la selección natural debido a que en su hábitat natural solamente existen semillas de tamaño pequeño y grande para las cuales los picos de longitud intermedia son menos aptos que los de longitud pequeña y grande, respectivamente. Es posible que si la selección disruptiva siguiese operando en un mismo sentido, es decir, favoreciendo los valores extremos del fenotipo en detrimento de los intermedios, y surgiese simultáneamente una preferencia en los pinzones cascanueces con distintas longitudes de pico por aparearse con los individuos de longitud de pico similar, podría acabar produciéndose un aislamiento reproductivo que fuera el precursor de la división de la especie en dos especies distintas.

.jpg){kind=link}
Por último, tenemos la selección equilibradora (traducida a veces como balanceada), en la cual ningún fenotipo tiene una ventaja mayor sobre el resto de fenotipos de la población, sino que existe un equilibrio entre todos ellos. Así, bajo un proceso de selección equilibradora, en una determinada circunstancia el fenotipo P podría tener ventaja sobre el fenotipo Q, mientras que en otra circunstancia podría suceder lo contrario, a saber, que Q tuviese una ventaja sobre P. La selección equilibradora contribuye de este modo al mantenimiento de la variedad genética de la población.
En función de las causas que están detrás de estos tipos de selección natural, los biólogos evolutivos distinguen entre selección ecológica y selección sexual. Todos los ejemplos que hemos expuesto en este artículo se refieren a la selección de tipo ecológico, en la que las presiones selectivas afectan a la habilidad de los individuos para escapar de los depredadores y conseguir alimento. Por su parte, en la selección sexual, las presiones selectivas que impulsan el cambio adaptativo en la población tienen que ver con cuestiones relacionadas con la reproducción, tales como la elección de pareja, la competencia intrasexual o la viabilidad de los gametos.
Para saber más:
Freeman, S., Quillin, K. & Allison, L. (2014). Biological Science (5th ed.). Londres: Pearson.
Lewontin, R. C. (1970). The Units of Selection. Annual Review of Ecology and Systematics, 1: 1-18.
Smith, T. B. (1987). Bill size polymorphism and intraspecific utilization in an African finch. Nature, 329: 717-719. Citado en: Freeman, S., Quillin, K. & Allison, L. (2014). Biological Science (5th ed.). Londres: Pearson.