

En temas anteriores hemos visto como la evolución afecta a diferentes caracteres del organismo, pero siempre hemos simplificado el proceso dando por hecho que el cambio genético y el carácter afectado es todo uno, sin intermediación de ningún tipo. La realidad es más compleja. Desde los genes que se han heredado durante la fecundación y que forman el genoma del cigoto hasta los diferentes caracteres y órganos que forman el organismo adulto, sujetos a los procesos de selección natural, hay un complejo proceso de desarrollo, sobre todo durante la etapa embrionaria, que en no pocas ocasiones es fundamental para entender cómo se ha producido la evolución. Al estudio evolutivo incorporando el desarrollo se le conoce coloquialmente como la EVO-DEVO, y ha recibido un impulso importantísimo en los últimos 30-40 años con el estudio en profundidad del desarrollo de varias especies modelo (Figura 1), entre las que podemos destacar: la mosca de la fruta Drosophila melanogaster, el pequeño gusano Caenorhabidtis elegans, el pez cebra Danio rerio, y el ratón (Mus musculus). A pesar de que el proceso de desarrollo presenta importantes diferencias en las 4 especies modelo, lo cierto es que la investigación en este campo ha demostrado que al igual que en otros aspectos de la biología, los procesos de desarrollo poseen altos niveles de conservación en muchos de sus procesos, lo cual ha servido para explicar cómo en numerosos casos la evolución se ha producido de una forma y no de otras. Como veremos, una de las grandes novedades que ha aportado este campo al estudio de la evolución es la posibilidad de que en muchos casos la evolución pueda estar constreñida/facilitada no solo por la mutación disponible sino también por el propio proceso de desarrollo.

Veamos un ejemplo de investigación sobre el desarrollo en un contexto evolutivo. Todos sabemos que los murciélagos son un grupo bastante especial dentro de los mamíferos, pues han desarrollado alas y unos sistemas de eco-localización muy especializados que les permiten orientarse en completa oscuridad. Estas novedades evolutivas han conseguido que este grupo de mamíferos sea uno de los más exitosos en términos evolutivos (representan el 20% de todas las especies de mamíferos). Los murciélagos poseen como parientes no alados más cercanos al grupo de las musarañas y en menor medida al de los ratones, que se han utilizado para comparar sus sistemas de desarrollo y así tratar de entender los procesos que explican su especial morfología. Así, por ejemplo, gracias a la anatomía comparada sabemos que las alas de los ratones no son más que manos que han evolucionado alargando sus falanges y extendiendo la piel que cubre las regiones entre las falanges. Desde hace años varios estudios comparativos entre el desarrollo en ratones y murciélagos han permitido empezar a desentrañar el tipo de cambios que la selección natural ha producido para favorecer la capacidad de vuelo en estos últimos. Se ha visto que la forma de la mano-ala de un murciélago se consigue manteniendo diferentes niveles de expresión en algunos morfogenes que determinan la proliferación celular de las células que van a dar lugar a las falanges y sus regiones adyacentes en ciertas fases del desarrollo. Así, por ejemplo, se han demostrado niveles más altos de expresión durante la morfogénesis de las alas en murciélagos comparada con la mano de ratones para los genes: HoxD, Tbx3 y la familia Fam, entre otros (Wang et al. 2014). La expresión de estos genes se puede estudiar durante la embriogénesis y visualizar su expresión con el objeto de mapear en que regiones se expresan y con qué intensidad, y de esa forma desentrañar cómo regulan el proceso de desarrollo que da lugar a las estructuras del adulto. De alguna forma este tipo de investigación nos permitirá entender cómo se produce la estructura y la forma de las alas en los murciélagos.
Mediante los diversos estudios de evo-devo se ha llegado a la conclusión de que existe un conjunto de características (herramientas) del desarrollo por las que la selección natural favorece o restringe el cambio de forma durante la evolución en cualquier grupo. Comentemos brevemente las cinco herramientas más importantes:
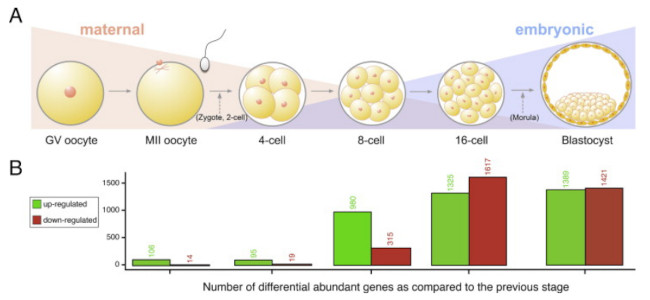
1. Procesos de activación e inactivación génica. El primer proceso que utiliza el ADN para regular el proceso de desarrollo es la activación/desactivación de diferentes genes a lo largo del desarrollo embrionario. No todos los genes se activan al mismo tiempo, ni en las mismas células que se van produciendo durante el desarrollo embrionario (ver ejemplo en Figura 2). En definitiva, buena parte de las diferencias de desarrollo se deben a la activación/desactivación diferencial de los mismos genes en las mismas o distintas células del embrión (Ejemplo en Graft et al. 2014).
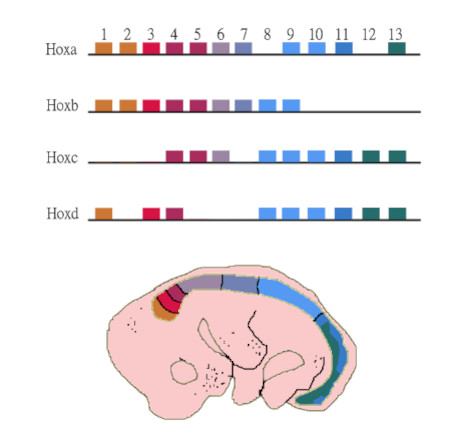
2. Compartimentalización y modularidad. Al estudiar de forma comparada el desarrollo de diferentes organismos se comprobó que todos tienen una estructura de genes reguladores muy particulares llamados los genes homeobox (por ejemplo, los genes HOX). Estos genes actúan compartimentalizando el eje antero-posterior de todos los organismos, y favoreciendo la diferenciación específica de los diferentes módulos corporales. Los genes Hox están muy extendidos en todos los animales y las diferentes compartimentalizaciones se han producido por duplicación génica y posterior cooptación de funciones (ver herramientas siguientes). Todos presentan una secuencia conservada característica y se encuentran ordenados en el organismo siguiendo el mismo orden de activación en el eje antero-posterior (ver Figura 3).
3. Cambios de ritmo durante la expresión. El desarrollo embrionario de dos organismos puede alcanzar a veces una considerable diferenciación simplemente cambiando el tiempo a la que acontecen los procesos de desarrollo (denominado heterocronía). Un ejemplo bien conocido es la evolución por neotenia del axolote mexicano (Figura 4). En el ambiente donde vive esta salamandra el presentar branquias (características de estadios larvarios) ha sido favorecido por selección natural. La forma en la que la selección natural ha favorecido dicho cambio ha sido mediante un cambio en los procesos de desarrollo de forma que las formas más juveniles de la especie han finalizado el desarrollo sin llegar a las formas adultas del resto de salamandras. La evolución del cráneo en nuestra propia especie también ha evolucionado mediante un proceso similar, pues el cráneo de Homo sapiens, se parece más a los cráneos de crías de otros primates que a la de los correspondientes adultos.

4. Duplicación y cooptación. Uno de los mecanismos más frecuentes de cambio evolutivo vía desarrollo es la duplicación génica, de forma que donde antes había un gen ahora hay dos o más. Las nuevas copias ya no son necesarias en la maquinaria bioquímica-morfogenética de las células y por lo tanto puede ser cooptado (utilizado en funciones nuevas gracias a la selección natural), lo que permite la aparición de innovaciones durante el desarrollo y la aparición de estructuras nuevas. Un ejemplo muy claro de cómo se ha producido este tipo de evolución lo tenemos en algunos estudios que han tratado de entender el paso de la unicelularidad a la multicelularidad, para ello una estrategia es el estudio de las volvocales, un grupo de organismos con especies unicelulares (Chlamydomonas spp) y pluricelulares de complejidad variable (Figura 5). En un estudio se comparó la expresión génica de una especie unicelular Chlamydomonas sp y otra pluricelular (Volvox sp para ver en cuantos genes difieran que afecten a la regulación de la mitosis (García-Fernández y Bueno 2020). Ambas especies comparten unos 14500 genes, pero solo difieren en un 0.22% que son específicos de Volvox. En particular los autores de este estudio encontraron que la especie multicelular presentaba 4 ciclinas D1 (molécula que regula el ciclo celular de la mitosis) comparadas con una sola en la unicelular. Estas cuatro ciclinas se han originado por duplicación génica de la ciclina original y en la especie multicelular han cooptado nuevas funciones para dar cuenta del proceso de replicación del nuevo organismo.

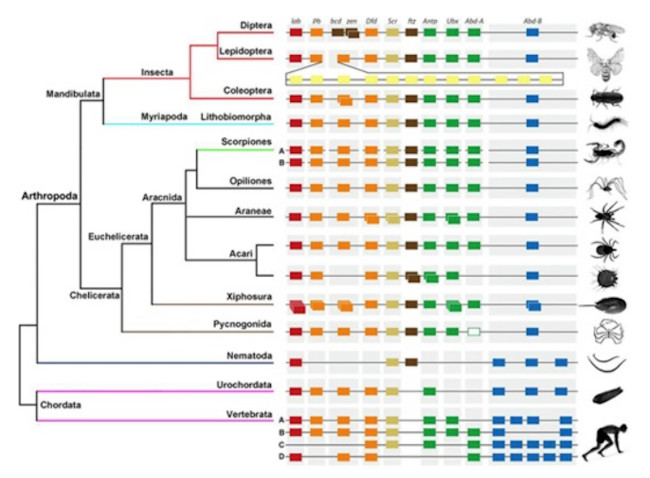
5. Alta conservación de los mecanismos anteriores. Todos los procesos anteriores se ha comprobado que se comparten en todos los organismos estudiados. Es más, en muchos casos las proteínas involucradas hacen las mismas funciones y presentan secuencias altamente conservadas, con lo cual podemos decir, que al igual que el código genético, lo fundamental de la maquinaria del desarrollo se mantiene muy conservado en los diferentes filos (Figura 6). Esto se ha comprobado al hacer inserciones de un morfogén (gen implicado en el desarrollo) de una especie en otra especie a menudo suele tener efectos modificadores del desarrollo. Es decir, cuando se inserta un morfo gen de la especie A en una especie B (pueden incluso ser relativamente distantes filogenéticamente), en la especie B se producen efectos similares a los que dicho morfogén realiza en la especie A.
También, podemos enfatizar que los procesos del desarrollo son responsables de la observación última de un fenómeno que hasta recientemente no se entendía bien del todo, la canalización evolutiva. Se denomina canalización a la constatación de límites en los procesos de desarrollo que hacen que procesos independientes de evolución acaben generando estructuras similares independientemente en dos especies. No hay que confundirlo con la convergencia evolutiva, donde la similitud sería sobre todo funcional y no estructural (alas de insecto y aves, por ejemplo) y sería dirigida en todo momento exclusivamente por la selección natural. Así, por ejemplo, se sabe que varios organismos muy diferentes han desarrollado sistemas de ecolocalización análogos (García-Fernández y Bueno 2020), como el que poseen delfines y murciélagos (estos últimos lo han generado al menos dos veces de forma independiente en dos linajes distintos). Un estudio detallado comparado de estos sistemas en murciélagos y delfines ha constatado una semejanza estructural muy grande, al estar formada por los mismos tipos celulares, afectados por las mismas proteínas (Prestina, Cadherina 23, Protocadherina 15, ortofelina, etc. En el caso concreto de la Prestina, proteína que forma parte de la estructura de las células ciliadas que detectan los ultrasonidos, es homóloga a la prestina que usan el resto de animales para poder oír el rango típico de sonido. Sin embargo, para que la prestina funcione con ultrasonidos, esta tiene que producir un cambio estructural que afecta a unos 20 aminoácidos. Curiosamente exactamente esos mismos 20 aminoácidos que han cambiado, lo han hecho de forma independiente en el linaje de murciélagos y delfines, y en 15 de esos aminoácidos se ha confirmado que son cambios no homólogos (García-Fernández y Bueno 2020). Esto demuestra que las posibilidades de generar células especializadas capaces de detectar y por lo tanto oír los ultrasonidos no son muy flexibles, y que la solución que adopta la selección natural en dos procesos independientes acaba llevando al mismo proceso de cambio concreto en la misma molécula, quizás porque otras alternativas o no son posibles o son muchísimo más improbables, este fenómeno es el que se conoce como canalización (constricción de la evolución como consecuencia del proceso de desarrollo). Podríamos decir que la canalización es el proceso contrario al de la plasticidad fenotípica (posibilidades de adaptación del proceso de desarrollo de un mismo genotipo a variantes ambientales).
Por último, otra de las constricciones mejor conocidas durante el desarrollo se da en la denominada constricción “bauplan” o de esquema corporal. Explica cosas que no se pueden hacer debido a la propia estructura corporal. Por ejemplo, los delfines adaptan el sistema pulmonar a la respiración en el agua, pero no generan branquias, porque tendrían que modificar su patrón de desarrollo embrionario temprano, lo cual es prácticamente imposible porque ya han adaptado un plan de desarrollo muy diferente. Es decir, las adaptaciones de organismos complejos normalmente se llevan a cabo en etapas más avanzadas del desarrollo embrionario. Esto es claro en cuanto que más temprano en la evolución, más se parecen los embriones, porque es más difícil que una mutación que afecte de forma importante a etapas tempranas del desarrollo no produzca un cambio catastrófico en el proceso de desarrollo posterior.
Para saber más:
Futuyma, D. 2009. Evolution. Second edition. Sinauer Associates, Sunderland.
García-Fernández, J., Bueno, D. 2020. El embrión inconformista. Como influye en nuestra evolución el desarrollo Embrionario. Editorial Universitat de Barcelona, Barcelona.
Graft et al. 2014. Genome activation in bovine embryos: a review of the literature and new insights from RNA sequencing experiments. Animal Reproduction Science. dx.doi.org/10.1016/j.anireprosci.2014.05.016.
Wang et al. 2014. Unique expression patterns of multiple key genes associated with the evolution of mammalian flight. Proceedings of the Royal Society, B, 281: 20133133.