

Una idea básica en evolución es que el ambiente es el motor del cambio causado por la selección natural. Sin embargo, el ambiente referido en términos evolutivos no sólo se refiere a las condiciones físicas del medio donde viven las especies, sino que de alguna forma también incluye todas las interacciones sociales que se pueden dar dentro de una especie (por ejemplo durante la interacción reproductiva), o incluso, como veremos en este tema, las interacciones con otras especies. Estos aspectos cobran especial importancia cuando la interacción evolutiva entre dos o más especies se mantiene estable en el tiempo, produciéndose una influencia continua y recíproca de una especie sobre la otra, afectando a sus respectivos procesos de evolución. Este fenómeno se conoce como coevolución. La coevolución suele tener dos acepciones que se pueden confundir, pero que se refieren a situaciones distintas en el contexto evolutivo (Futuyma 2009): coevolución difusa y coevolución estricta.
Coevolución difusa
La coevolución difusa, se aplica cuando consideramos que todas las especies que viven de un determinado ecosistema modifican de alguna manera el medio ambiente del ecosistema, y que por tanto influyen a cualquier especie presente en dicho ecosistema.
Coevolución estricta
La coevolución estricta se da cuando se puede demostrar que la evolución de una especie influye en la evolución de otra, y esto también ocurre recíprocamente. La parte más delicada aquí es el término “se puede demostrar” porque como veremos, lograr evidencias de esto último no es tan fácil, e incluso en los casos mejor documentados persiste la duda de cómo interpretarlos, veamos un ejemplo.
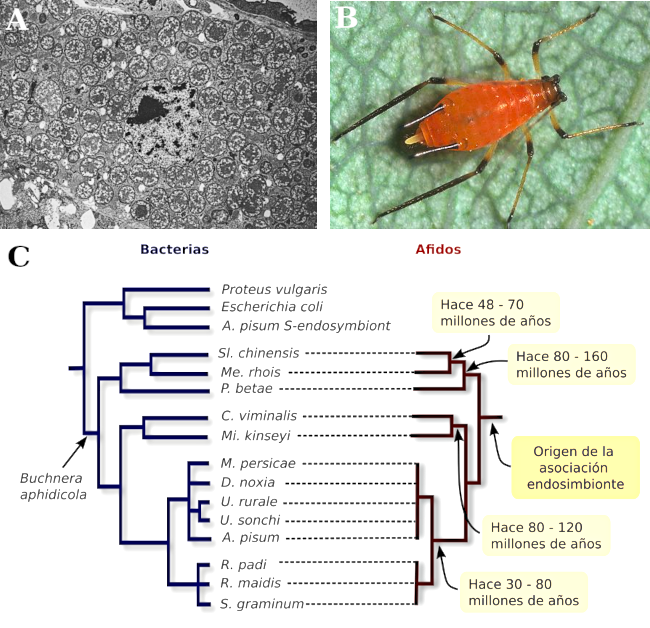
Los áfidos son un grupo de insectos que se alimentan de la savia de las plantas, algunas de sus especies son conocidas pestes de muchas especies agrícolas. Los áfidos viven en una asociación obligada desde hace unos 100 millones de años con un grupo de bacterias endosimbiontes (viven dentro de algunas células del insecto) del género Buchnera (género filogenéticamente emparentado con Escherichia coli) que viven en sus tractos digestivos. Estas bacterias permiten a los áfidos conseguir ciertos aminoácidos y vitaminas que de otra forma no lograrían con su dieta de savia vegetal (Figura 1). Cuando se compara la filogenia de las especies de áfido y la de su endosimbionte se observa una concordancia perfecta en los procesos de diferenciación y especiación, lo que sugiere una estrecha coevolución (Figura 1c). No obstante, esta misma filogenia puede servir para rechazar la hipótesis de una coevolución estricta. Así, otros autores postulan que se ha producido un proceso de especiación del género de áfidos, y que su hospedador simplemente ha tenido que seguir ese proceso de forma obligada al ser un endosimbionte. Es decir, aceptan la influencia de los áfidos en la evolución de su endosimbionte pero niegan que sea recíproca.
{kind=link}
{kind=link}
Interacciones sin y con coevolución
Las interacciones entre especies mediadas por la selección natural pueden ser muy diversas, y no todas son estrictamente coevolución. Por ejemplo, algunas especies han evolucionado para presentar un fenotipo que se camufla con su entorno (Figura 2). En este caso es obvio que hay una interacción entre especies (animal y vegetal), mediada por selección pero sin coevolución. El camuflaje puede evolucionar en un contexto de especies donde dicho carácter les ayuda a escapar de sus depredadores pero también en algunos depredadores para que no los vean sus presas.

{kind=link}
_male,_nymph,_Arnhem,_the_Netherlands.jpg){kind=link}
{kind=link}
{kind=link}
Otro ejemplo bastante espectacular de interacción entre especies (aunque no implique necesariamente coevolución) mediada por selección natural es la coloración aposemática (coloración vistosa, coloración que ha evolucionado para que otras especies la reconozcan como peligrosa o no comestible y la ignoren; Figura 3). La coloración aposemática ha evolucionado porque las especies venenosas o de mal sabor pueden favorecer la evolución de colores muy vistosos si estos facilitan que los ejemplares que los portan sean atacados en menor frecuencia por posibles depredadores, respecto a ejemplares que no porten dicha coloración.

{kind=link}
{kind=link}
Por último podemos mencionar los fenotipos de imitación (miméticos; en este caso sí suele implicar coevolución). La coloración de imitación se da habitualmente cuando una especie evoluciona para presentar las formas y colores de otra. Se conocen dos situaciones con el nombre de mimetismo mülleriano y batesiano, en honor a sus respectivos descubridores (Futuyma 2009). El mimetismo mülleriano se da cuando dos o más especies (todas ellas igualmente peligrosas o de mal sabor) producen un mismo patrón de coloración, y la idea es que la selección natural favorece cualquier mutación coincidente entre ellas porque de esa forma la señal de aviso será más eficaz. Por último el mimetismo batesiano se daría cuando sólo una de las especies es la que da una señal auténtica de peligro, mientras que la otra ha copiado el mismo patrón que la peligrosa pero sin serlo (por ejemplo la falsa coral en la Figura 4 que imita el patrón de color de la auténticamente venenosa de la Figura 3c).

{kind=link}
Los casos de coevolución más estudiados los veremos en los próximos dos apartados, subdivididos como coevolución negativa o positiva.
Para saber más:
Futuyma, D. 2009. Evolution, second edition. Sinauer Associates, Inc., Massachusetts.
Moran, N. y P. Boumann. 1994. Phylogenetics of cytoplasmically inherited microorganisms of arthropods. Trends in Ecology and Evolution, 9: 15-20.