

Las primeras células que aparecieron en la Tierra fueron las células procariotas hace unos 3500 millones de años. Procariota significa anterior al núcleo, es decir, no tienen el ADN encerrado en un compartimento membranoso. De hecho, estas células tienen una organización relativamente sencilla con una membrana que delimita un espacio interno donde se producen las reacciones químicas. Por fuera de la membrana tienen una pared celular, una cápsula y en ocasiones muestran prolongaciones como son los flagelos bacterianos que permiten la movilidad, y los pilis para el intercambio de material genético. Esta forma celular fue la única en los primeros cientos de millones de años de la vida en la Tierra. Se conocen dos grandes grupos de procariotas: las bacterias y las arqueas.
Al proceso de formación de las células eucariotas a partir de las procariotas se llama eucariogénesis y supuso una transición evolutiva, es decir, se formó un tipo celular nuevo y diferente a lo que había anteriormente, presentando suficientes novedades como para abrir nuevos caminos evolutivos hasta entonces inexplorados. Así, las células eucariotas llegaron a una complejidad morfológica y estructural no conocida hasta entonces, destacando un complejo sistema de compartimentos membranosos internos, incluido el núcleo, organizados por un citoesqueleto, y aumentaron enormemente la cantidad de ADN por célula en comparación con los procariotas. También fueron capaces de incorporar genomas completos de otras células (que dieron lugar a las mitocondrias y a los cloroplastos), descubrieron la reproducción sexual, y permitieron la aparición de algo desconocido hasta entonces: los organismos pluricelulares (cosa que ha ocurrido varias veces de forma independiente).
1. ¿Cuándo?
La aparición de la célula eucariota fue un evento evolutivo que ocurrió hace unos 1500-2000 millones de años, es decir, unos 1500 millones de años después de que lo hicieran las primeras células procariotas. Esto implica que el proceso de formación de las células eucariotas, o eucariogénesis, comenzó mucho antes, quizá en las comunidades de procariotas que habitaron la Tierra hace entre 2500 y 4000 millones de años.
2. ¿De quién?
Esta cuestión no está resuelta todavía, pero se acepta que fue la consecuencia de la colaboración entre los dos tipos celulares procariotas que existían entonces: las arqueas y las bacterias. Se propone que las eucariotas son monofiléticas, es decir, todas las células eucariotas, incluyendo plantas, animales, hongos, algas y las eucariotas unicelulares, descienden de un único ancestro denominado LECA (last eukaryotic common ancestor). Por tanto, y mientras no se demuestre lo contrario, la célula eucariota sólo se inventó una vez durante la evolución. Mediante el estudio comparado de genes se ha llegado a la conclusión de que LECA tenía un genoma tan complejo como los eucariotas actuales y probablemente era morfológica y estructuralmente parecida a los eucariotas actuales..
No cabe duda de que LECA se formó a partir de células procariotas, pero ¿a partir de cuál se formó, de las bacterias o de las arqueas? Hay un gran problema en esta elección. Las células eucariotas actuales parecen ser quimeras, en las que coexisten genes heredados de los dos tipos de procariotas. Durante la eucariogénesis se formó un genoma con genes de origen arqueano, bacteriano por transferencia horizontal, y también se incorporaron genes provenientes del endosimbionte, una bacteria, cuyo descendiente actual son las mitocondrias. Hay una gran división del trabajo entre los genes arqueanos y los bacterianos. Los genes arqueanos trabajan en la traducción, transcripción y replicación del ADN, mientras que los bacterianos están relacionados con el metabolismo, biosíntesis de enzimas, glucólisis y síntesis de aminoácidos. Sin embargo, hay algunos genes que escapan a esta regla.
Para complicar más la cosa, aquellos genes de origen bacteriano, y algunos arqueanos, no proceden de un solo grupo, sino que son el legado de varios grupos. A pesar de toda esta complejidad, numerosas evidencias tienden a colocar a los eucariotas en la rama de las arqueas (ver Figura 1).

Atendiendo a los estudios filogenéticos (comparación de secuencias de nucleótidos de algunos genes) se tiende a colocar actualmente a los eucariotas como descendientes del grupo de las arqueas denominadas Asgard, entre las que están las lokiarqueas. Las lokiarqueas son evolutivamente las procariotas más próximas a los eucariotas. Hay que tener cuidado con estas clasificaciones porque para ellas se utilizan los genes denominados informacionales (aquellos encargados de procesar el ADN y que se supone que han cambiado menos a lo largo del tiempo). Los que defienden este origen consideran que estos genes son los que se transmiten de “padres” a “hijos” directamente y no entre células no relacionadas, y por tanto los importantes a la hora de establecer relaciones evolutivas. Además, y esto es importante, en algunos miembros de la familia Asgard, incluyendo a las lokiarqueas, se han encontrado genes de proteínas que se creían exclusivas de eucariotas, tales como los relacionadas con el citoesqueleto, con la organización de las membranas internas, con el tráfico vesicular y con la reparación del ADN, y que no existen conjuntamente en ningún otro grupo de procariotas.
Curiosamente, la existencia de las lokiarqueas se ha deducido por estudios de metagenómica, es decir, se cogió agua cerca de una fumarola, se aisló todo el ADN que contenía, se ensamblaron en genomas, y posteriormente se determinó cuántas células diferentes se podían obtener de tales genomas. Las arqueas Asgard se han encontrado en lugares anóxicos, lo que hace presuponer que las células ancestrales vivían en ambientes sin oxígeno y que sólo gracias a la adquisición de las bacterias tolerantes al oxígeno (las futuras mitocondrias) pudieron colonizar ambientes más oxigenados. Al no saber cómo son morfológicamente no se pueden establecer similitudes con las células eucariotas en cuanto a tamaño o complejidad estructural.
De cualquier forma es más complicado de lo que parece, y hay que integrar información adicional en los árboles filogenéticos. Por ejemplo, en el núcleo de una eucariota hay 2 o 3 veces más genes de origen bacteriano que de arqueas. Estos genes parecen provenir de muchas fuentes tales como cianobacterias, actinobacterias, proteobacterias y otras. Hay otro problema adicional, las membranas de los eucariotas no tienen cadenas de isoprenoides en sus ácidos grasos ni enlaces tipo éter, ambos típicos de las membranas de las arqueas, y por tanto se puede decir que tienen una membrana más bacteriana. Por otro lado, hay una gran cantidad de genes bacterianos en las eucariotas que indican el gran papel que tuvieron las bacterias durante la eucariogénesis. Esto debería considerarse cuando se representa a las eucariotas en el árbol de la vida.
3. ¿Cómo? Eucariogénesis
Hay un hecho clave en la aparición de LECA y es qué importancia tuvo la incorporación del antepasado de las mitocondrias, es decir, cuando se produjo la endosimbiosis. Hay autores que sugieren que esta incorporación fue la desencadenante y motor de la evolución hasta LECA, mientras que otros autores sugieren que la célula que “engulló” al ancestro bacteriano de las mitocondrias ya era muy compleja, tanto genómicamente como estructuralmente, y por tanto la endosimbiosis sólo fue un paso más en la evolución hasta LECA (ver Figura 2). Hay multitud de modelos que intentan explicar cómo ocurrió el proceso evolutivo que desembocó en LECA, pero hay dos líneas principales:
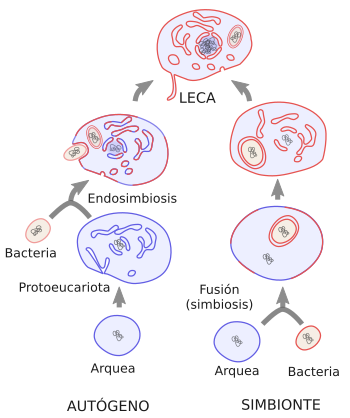
Modelo autógeno o del eocito. La eucariogénesis fue un proceso progresivo de adquisición de la complejidad celular que siguió estos pasos: arquea - FECA (first eukaryotic common ancestor) - LECA. Existió una célula protoeucariota (FECA) que procedió de un ancestro arqueano (del grupo Asgard). Esta célula evolucionó de manera independiente, adquiriendo la mayoría de las complejidades que aparecen en una célula eucariota actual, incluyendo endomembranas y citoesqueleto, pero aún no tendría a las mitocondrias. Tuvo la capacidad de fagocitar y en algún momento incorporó a una bacteria ancestral a las alfa-protobacterias actuales, que no fue digerida, sino que pasó a vivir como endosimbionte. Este modelo está recibiendo cada vez más soporte porque a medida que se van descubriendo nuevos genomas arqueanos se está encontrando que los genes responsables del citoesqueleto y el tráfico de membranas ya estaban presentes en la rama de la arquea de la cual se propone que surgieron los eucariotas, por lo que una evolución progresiva sería posible. La complejidad genómica se iría incrementando con duplicaciones génicas y adquisición por transferencia horizontal de nuevos genes durante la transformación de FECA en LECA. Hay algunas evidencias que indican que la tasa de duplicaciones génicas aumentó tras el proceso de endosimbiosis. Sin embargo, algo a lo que tiene que responder este modelo es por qué no se han encontrado células eucariotas sin mitocondrias (aquellas células actuales que no tienen mitocondrias tienen otros orgánulos derivados de éstas).
Modelo simbionte. Propone una fusión directa entre una arquea y una bacteria, y no existiría un protoeucariota como tal. Aquí habría sólo dos ramas principales de células iniciales, arqueas y bacterias. Los eucariotas serían una tercera rama surgida de la fusión de estas dos ramas. Este evento de fusión disparó el proceso de incremento de complejidad celular, y la bacteria terminaría siendo una mitocondria. En esta simbiosis las dos células se repartieron el funcionamiento celular: arquea el ADN y bacteria el metabolismo. Hay una variante de este modelo en la que la asociación entre bacterias y arqueas no tuvo por qué ser una incorporación de una célula dentro de otra en un momento determinado, sino que la asociación ocurrió a lo largo de mucho tiempo. Habría ocurrido transferencia lateral de genes de la bacteria a la arquea debido a que las condiciones ambientales favorecieron la proximidad física entre ambas. Se propone la teoría del hidrógeno en el que la bacteria produciría hidrógeno para el metabolismo de la arquea y la arquea produciría sustancias carbonadas que usaría la bacteria. Finalmente hubo una incorporación física de la bacteria dentro de la arquea, la cual ya tenía muchos genes bacterianos. Cómo se produjo esta inclusión no está claro. Se ha asumido que fue por fagocitosis, pero en realidad no hay ninguna evidencia experimental que apoye esta idea.
4. Endomembranas
Una de las características que distingue a las células eucariotas de las procariotas es su sistema de endomembranas organizado en compartimentos llamados orgánulos, cuyo origen evolutivo todavía permanece sin aclarar. Se proponen dos procesos: a) los procariotas actuales no generan vesículas internas, es decir, no son capaces de hacer endocitosis, aunque sí pueden invaginar su membrana y crear cisternas membranosas hacia el interior que permanecen conectadas con la membrana plasmática. Estas cisternas se propone que se separaron creando los orgánulos internos. Como se ha mencionado anteriormente, el descubrimiento de proteínas relacionadas con el tráfico vesicular, la organización membranosa y el citoesqueleto en grupos nuevos de arqueas indica que la posibilidad de una organización compleja interna podría haberse generado ya en la arqueas, antes de la llegada de las mitocondrias. b) Sin embargo, tanto arqueas como bacterias pueden generar vesículas hacia el exterior, es decir, vesículas extracelulares. Es muy interesante que esta capacidad se mantiene en las mitocondrias actuales, las cuales son capaces de generar vesículas denominadas "vesículas derivadas de mitocondrias". Estas vesículas quedan libres en el citosol y, durante la eucariogénesis, pudieron fusionarse y organizarse en compartimentos.
Aceptando que la célula hospedadora fue un ancestro arqueano y la huésped fue un ancestro bacteriano, permanece la cuestión de cómo se cambiaron los lípidos de la membrana plasmática de origen arqueano (cadenas de isoprenoides y enlaces éter en los lípidos) por los de origen bacteriano actuales (lípidos sin isoprenos y sin enlaces tipo éster).
Para saber más:
Archivald, JM. 2015. Endosymbiosis and eukaryotic cell evolution. Current biology. 25: R911-R921.
López-García P, Moreira D. 2015. Open questions on the origin of eukaryotes. Trends in ecology and evolution. 30: 697-708.
Nobs S-J, MacLeod FI. Wong HL, Burns BP. 2022. Eukarya the chimera: eukaryotes, a secondary innovation of the two domains of life? Trends in microbiology. 30: 421-431.